Impact of Mineral and Vitamin Status on Beef Cattle Immune Function and Health
![]()
Supplementing Trace Minerals to Beef Cows during Gestation to Raise Productive and Health Responses of the Offspring
one
Prairie Research Unit, Mississippi State University, Prairie, MS 39756, United states of america
2
Department of Animal Science, Texas A&M University, Higher Station, TX 77843, USA
3
Department of Animate being and Range Sciences, Montana State Academy, Bozeman, MT 59717, USA
*
Writer to whom correspondence should exist addressed.
Academic Editors: Mandy Paterson and Alejandro East. Relling
Received: 4 March 2021 / Revised: xiv April 2021 / Accepted: 15 April 2021 / Published: 18 April 2021
Simple Summary
During gestation, the fetus relies on the dam for the supply of all nutrients, including trace minerals, which are essential for developmental processes including organogenesis, vascularization, and differentiation. Alterations in maternal nutritional condition may promote adaptations that permanently alter the trajectory of growth, physiology, and metabolism of the offspring. Supplementing trace minerals to gestating cows may exist a strategy to enhance progeny operation and health. The purpose of this review is to highlight current information relevant to trace mineral supplementation during gestation, with an emphasis on Zn, Cu, Co, and Mn, and their impacts on offspring productive responses. Identifying nutritional strategies targeted at this catamenia of evolution and understanding the implications of such provides an opportunity to raise the productive efficiency of beef cattle systems.
Abstract
Nutritional management during gestation is critical to optimize the efficiency and profitability of beefiness production systems. Given the essentiality of trace minerals to fetal developmental processes, their supplementation represents one approach to optimize offspring productivity. Our research group investigated the impacts of supplementing gestating beef cows with organic-complexed (AAC) or inorganic sources (INR) of Co, Cu, Mn, or Zn on productive and health responses of the progeny. Calves born to AAC supplemented cows had reduced incidence of bovine respiratory disease and were >xx kg heavier from weaning until slaughter compared to unsupplemented cohorts. Complementing these findings, heifer progeny born to AAC supplemented cows had accelerated puberty attainment. Collectively, research demonstrates supplementing trace minerals to gestating beef cows may exist a strategy to enhance offspring productivity in beef production systems.
i. Introduction
Over the course of the next xxx years, the global population is expected to reach over 9.1 billion people, and as a result, nutrient production volition demand to increase by over 70% [ane]. Of the total increment in food production required, meat product will need to rise past approximately 200 million tonnes and accordingly, in that location will be greater demand for beef products [1]. Concomitantly, urban areas volition expand, reducing natural resource and land available for beef cattle production and other agricultural activities. Hence, advancements in both management and technologies are required to reach a level of beast production that tin provide for the population by 2050 [1].
Moo-cow-calf production systems are the foundation for the U.s.a. and worldwide beef industries by determining the number of cattle available for harvest. The success of each cow-calf operation depends on its ability to produce one healthy calf per cow annually, and cow-calf producers are constantly challenged to better growth, efficiency, carcass, musculus, and quality characteristics of offspring [2]. Nutritional manipulation during periods of developmental plasticity such every bit the embryonic, fetal, and neonatal periods exerts short- and long-term consequences on skeletal muscle and adipose tissue development, health, and overall performance of offspring [3,iv]. Improved skeletal musculus development and adipogenesis in skeletal musculus raise growth performance and carcass marbling upon slaughter [5], traits that benefit carcass value and beef palatability [6]. Moreover, the development of replacement beef heifers is a critical component of cow-calf systems, and production efficiency increases with improved female person longevity [seven]. Maternal nutrient status during fetal development also affects the lifetime productivity of female person progeny by impacting ovarian reserves [8,ix]. Identifying specific nutritional strategies that are targeted at these disquisitional periods of evolution provides a unique opportunity to optimize the efficiency and profitability of beef cattle systems. Hence, the purpose of this review is to highlight current data relevant to trace mineral supplementation to beef cows during gestation, with an accent on Zn, Cu, Co, and Mn, and their impacts on offspring productive responses.
ii. Developmental Programming
Developmental programming, also chosen fetal programming, proposes that alterations in fetal nutritional or endocrine status may result in adaptations that permanently alter the trajectory of growth, physiology, and metabolism of the offspring [iii,10]. Moreover, recent reviews focused on developmental programming have suggested that nutritional manipulation during gestation can have profound influences on the occurrences of adulthood affliction and long-term operation [iii,11], given that development of the fetus is sensitive to direct and indirect effects of maternal nutrition at all stages between oocyte maturation to parturition [12,13]. Hence, effective prenatal nutritional management represents an opportunity to alter offspring evolution and performance.
The fetal stage is disquisitional for skeletal muscle and intramuscular adipocyte development [14,15], whereas a reduction in the number of musculus fibers permanently reduces animal operation, and adipocyte development provides sites for marbling formation during the feedlot finishing menses [five]. Maternal nutrient restriction results in a reduction in the full number of secondary muscle fibers [xiv], however, maternal overnutrition results in elevated expression of adipogenic genes in fetal skeletal muscle [sixteen] and increased number and size of adipocytes in the skeletal musculus postnatally [17]. The effect of maternal food condition during gestation can also have considerable impacts on the future reproductive functioning of female progeny. For heifers to be efficient, they need to attain puberty by 12 months of historic period and conceive early in their first breeding flavour [18]. Therefore, nutritional management to maximize the number of replacement heifers pubertal by 12 months of age are critical to the productivity of moo-cow-calf operations. For example, heifers built-in to protein supplemented dams had greater conception rates during the get-go 21 days of the convenance season [nineteen]. Heifers born to energy-restricted dams had a reduction in antral follicle count compared to heifers built-in to control-fed cohorts [9,20,21], a trait that is positively correlated with pregnancy success in both beef heifers and dairy cows [22,23,24]. Suboptimal maternal food status during gestation as well results in alterations within the neuroendocrine system of the female offspring, leading to delayed puberty attainment and disrupted reproductive function [25]. Taken together, these findings betoken maternal nutritional condition during gestation significantly impacts offspring growth and development [three], leading to long-term furnishings on functioning and quality of carcass traits [3,26] and altered reproductive function of female progeny [27].
2.1. Dietary Factors and Developmental Programming
Maternal macronutrient nutrition has been reviewed several times in the literature [three,xiii,28], just little is known about the furnishings of maternal trace mineral supplementation on progeny wellness and operation. For example, protein supplementation provided to cows during late gestation has been shown to increment offspring birth and weaning weights [29], increase the postweaning charge per unit of bodyweight proceeds [19], and enhance carcass marbling [26]. Increasing dietary energy during late gestation results in increased progeny birth torso weight [30], whereas energy source during gestation too affects fetal growth. Research has demonstrated offspring from cows supplemented with corn or dried distillers' grains with solubles were heavier at birth and tended to be heavier at weaning compared with offspring from cows fed grass hay at isocaloric intake [31]. Other nutrients, including trace minerals, are essential for fetal developmental processes including protein synthesis, os formation, and lipid metabolism [32], and warranted investigation.
two.ii. Trace Minerals
Cow-calf production systems depend on forages as the main source of nutrients to cattle, which may be variable in quality and might not be nutritionally complete [33]. Forages provide minerals that are vital for nigh all processes inside the body. Many factors affect the mineral content of forages including soil, plant species, stage of maturity, yield, climate, and direction practices [33]. However, forages often practise not provide enough minerals to run across the requirements of livestock [34]. Hence, moo-cow-calf producers must design and integrate mineral supplementation programs according to animal requirements and fodder characteristics to maintain maximum long-term productivity [35]. Minerals are divided into 2 classes; macro minerals which are required in amounts > 100 ppm of the nutrition, and trace minerals which are required in amounts < 100 ppm of the diet and do not alter with animal stage or level of product [36]. Although trace minerals such as Cu, Zn, I, Mn, Se, Co, and Atomic number 26 are present in only minute amounts, they are essential for beef cattle homeostasis [36]. In fact, trace mineral deficiencies during fetal life are known to persistently touch on the growth, immunological, and morphological evolution of a variety of fetal and neonatal tissues [37] given that these elements play numerous roles in enzymatic and metabolic functions during fetal evolution [32]. Still, the fetus relies fully on the maternal–fetal interface for a proper supply of trace minerals [38]. For example, Gooneratne and Christensen [39] reported that fetal liver Cu concentrations are greater than that of the maternal liver, suggesting that the maternal system shunts Cu to back up fetal development [39]. Hansard [forty] investigated the placental transfer of radioactive Mn from maternal to fetal tissue and noted that only xv% of the radioactive Mn was absorbed by the cows and 70% of the Mn was deposited in fetal tissue. Hence, these results signal that fetal requirements for trace minerals accept priority over maternal requirements for these elements throughout gestation.
ii.iii. Inorganic vs. Organic Trace Mineral Supplementation
Trace minerals have been traditionally added to brute diets to meet requirements, however, the source of mineral supplementation may bear on individual animal functioning. Organic trace minerals differ from inorganic sources due to their chemical association with an organic ligand, as normally trace minerals are found as inorganic salts [41]. Recently, there has been marked interest in the use of organic trace minerals in ruminant diets due to indications that these may be of greater bioavailability compared to their inorganic counterparts [42]. Metallic chelates, or complexes that constitute an organically bound trace mineral, are hypothesized to be stable in the digestive tract, thus protecting the metal from forming complexes with other dietary components that would inhibit assimilation, and perhaps allowing greater absorption of the trace mineral [41]. In social club for an animal to utilize inorganic trace minerals, the fauna must outset convert them to organic biologically active forms, given that trace minerals in the torso function well-nigh entirely every bit organic complexes or chelates already [41]. Therefore, ruminants may reply to supplementation in the grade of enhanced growth, milk product, reproduction, and immune responses.
Supporting this rationale, Nocek et al. [43] reported that cows can be supplemented with organic trace minerals at 75% of requirements with no reduction in reproductive or productive performance compared with supplementing 100% of requirements using simply sulfate sources. Organic trace mineral supplementation to dairy cows significantly increased milk production, milk fat yield, and milk protein yield compared to inorganic trace mineral supplemented cohorts [44]. Previous research has demonstrated supplemented organic complexed trace minerals improved reproductive performance in cattle, decreasing interval to starting time estrus and increasing pregnancy rates [45,46], as well as resulting in improved in vitro embryo production efficiency [47]. A meta-analysis by Rabiee et al. [48] reported that lactating dairy cows supplemented with organic-complexed trace minerals take greater milk yield, fewer days open, and crave fewer services per conception compared with inorganic supplemented cohorts. Ahola et al. [49] also reported an improved pregnancy rate to AI of grazing beefiness cows supplemented with organic-complexed trace minerals compared with not-supplemented cohorts. Collectively, these results demonstrate organic-complexed trace mineral supplementation positively impacts cow productive and reproductive responses.
Supplying organic complexed trace minerals has as well been shown to enhance creature productivity and immune parameters during challenging stages such as the periparturient flow [50,51], peradventure due to the participation of trace elements in enzymatic and antioxidant systems [52,53] that may take major implications to future progeny operation. In fact, organic Cu, Mn, and Zn supplementation during late gestation improved calf serum antioxidant capacity and immunoglobulin concentrations compared with calves from non-supplemented cows [54]. Formigoni et al. [55] supplemented inorganic or organic Zn, Mn, and Cu to gestating Holstein cows and noted an increased concentration of immunoglobulins in the colostrum and reduced dogie mortality in cows fed organic trace minerals compared with inorganic-supplemented cohorts. Still, research investigating the impacts of trace mineral source supplementation on beef cattle performance and health has been variable, potentially due to interacting factors nowadays across experiments including, just not limited to, trace mineral antagonists, environmental pressures, duration of supplementation, and maternal trace mineral condition [33,49,56].
3. Supplementing Organic-Complexed Trace Minerals during Late-Gestation
The fetus depends on the dam for the proper supply of all nutrients during gestation, including trace minerals [38]. Further, if maternal supply is inadequate or supply to the fetus is impaired, developmental processes and postnatal functioning may be hindered [57]. Trace elements such as Zn, Cu, Mn, and Co are required for the proper evolution of fetal nervous, reproductive, and immune systems [32,58], underscoring the importance of supplying these nutrients to gestating beef cattle. Hostetler et al. [32] reported supplementing an organic Cu, Mn, and Zn to gestating sows increased concentrations of these trace minerals in fetal tissues and reduced fetal loss by d 30 of gestation. Supporting this rationale, our research grouping reported that supplementing late gestating beef cows with an organic complexed source of Co, Cu, Zn, and Mn instead of no supplementation optimized offspring productivity [59]. This initial project was conducted with beefiness cows supplemented with sulfate sources of Cu, Co, Mn, and Zn (INR), organic complexed Cu, Co, Mn, and Zn (AAC), or no supplemental Cu, Co, Mn, and Zn. Marques et al. [59] conducted this trial using cows at the end of their 2d trimester of gestation (d 0 of the experiment), and formulated all diets to meet the requirements for free energy, protein, Se, I, and vitamins of meaning cows during the last trimester of gestation [60]. The INR and AAC sources were formulated to provide the same daily amount of Cu, Co, Mn, and Zn. As expected, supplementing INR or AAC increased moo-cow liver concentration of Co, Cu, and Zn in samples collected prior to calving (d 75) compared with non-supplemented cohorts (Table 1). Marques et al. [59] also reported liver Cu and Zn concentrations in the neonatal calf were only increased in calves born to AAC supplemented cows compared with not-supplemented cows (Table 1). As stated previously, organic trace minerals are expected to have enhanced absorption, retention, and overall bioavailability [32,41], nevertheless only moo-cow liver Co concentrations supported this rationale. Corroborating the results of Marques et al. [59] others accept also reported variable effects of supplementing organic Zn, Cu, and Co on the liver mineral status of beefiness cattle [49,61,62].
The current requirements for trace minerals were established to support normal growth, reproductive, and immune functions but appear to exist insufficient when cattle are exposed to stressors from management practices, such every bit weaning, road ship, and arrival at the feedlot [63]. Given the essentiality of Cu, Mn, Co, and Zn for proper evolution of the fetal immune system [31,48] and their profound effects on antioxidant and enzyme components [64,65], maternal dietary trace mineral supply might be 1 feasible nutritional alternative to program the allowed system of the fetus. Accordingly, Jacometo et al. [66] reported that calves born from cows supplemented with organic trace minerals during belatedly gestation had decreased expression of markers of inflammation and oxidative stress at three weeks of historic period compared to calves born to inorganic supplemented cows. These authors hypothesized that maternal supplementation with organic trace minerals, may alter the immune response of the neonate, only noted that further research was needed to make up one's mind if the effects would persist [66]. In turn, Marques et al. [59] reported that calves born to AAC cows had a reduced BRD incidence compared with calves from not-supplemented cows (Table i), suggesting that feeding the AAC diet to belatedly gestating beef cows resulted in programming furnishings on postnatal offspring health.
Equally previously mentioned, maternal nutrient inputs during gestation have a large impact on biological mechanisms involved in fetal growth and nutrient utilization that may impact progeny performance and health afterward in life [67,68]. The fetal growth trajectory is persistently afflicted by maternal food intake from the early stages of fetal development to birth, whereas the majority of research is directed toward the last trimester of gestation when approximately 75% of the fetal growth occurs and nutrient requirements for fetal growth are maximal [36,68]. Trace minerals, for case, are involved in all stages of prison cell growth and differentiation and are vital components of many enzymes and cell structures [69]. These elements affect fetal development by altering hormones, growth factors, and jail cell signaling pathways involved in food uptake by the fetus, which may irreversibly impact progeny productivity [3,69,70]. Although no treatment differences were detected for calf birth body weight (BW), Marques et al. [59] reported that organic Cu, Co, Mn, and Zn supplementation to beef cows during tardily-gestation increased BW of their offspring by 24 kg at weaning and past 31 kg at slaughter compared with calves from non-supplemented cows (Table 1). Others investigating trace mineral supply in maternal diets have also demonstrated changes in progeny operation trajectory [49,61]. Stanton et al. [61] noted that supplementing tardily-gestating beefiness cows with organic-complexed Cu, Zn, Co, and Mn improved progeny weaning weight compared with calves from cows that received sulfate sources. Collectively, the results from Marques et al. [59] were novel, and suggestive of programming furnishings of organic trace minerals on postnatal offspring performance and health [13]. Notwithstanding, Marques et al. [59] recognized that the physiological mechanisms underlying these outcomes were nevertheless unknown and warranted investigation.
4. Supplementing Organic-Complexed Trace Minerals Mid- to Late-Gestation
Nutritional direction of beef cows impacts fetal development throughout the entirety of gestation [3], whereas Marques et al. [59] investigated supplementation during the last trimester of gestation. Supplementing organic trace minerals may be of even greater benefit if offered to beefiness cows over a greater duration of gestation. Moreover, Marques et al. [59] did not evaluate the potential physiological mechanisms by which organic trace mineral supplementation positively impacted postnatal offspring performance and wellness [59]. A majority of U.Southward. cow-calf herds receive supplementation [71] underscoring the importance of deciphering the difference between organic complexed and sulfate sources of trace minerals, without the inclusion of a not-supplemented group. Therefore, the next step was to evaluate the effects of supplemental organic complexed or sulfate sources of Co, Cu, Mn, and Zn to beef cows during the second and third trimesters of gestation.
In Harvey et al. [72], non-lactating meaning beef cows were supplemented with sulfate sources of Cu, Co, Mn, and Zn (INR) or organic complexed Cu, Co, Mn, and Zn (AAC) outset at the finish of the get-go trimester of gestation (d 0 of the experiment). The INR and AAC diets were formulated to provide the same daily amount of energy, protein, macro minerals, and trace minerals including Co, Cu, Mn, and Zn [59]. In samples collected during late gestation (d 97), Harvey et al. [72] reported greater liver Co, less liver Cu, and similar liver Zn and Mn between cows supplemented with sulfate or organic complexed sources (Table 2), corroborating results reported by Marques et al. [59]. Even so, Harvey et al. [72] reported greater liver Cu and Zn concentrations in cows receiving INR diets compared to AAC cohorts upon calving, whereas no handling differences were detected for liver Co or Mn (Table 2). These latter outcomes do non corroborate that organic trace mineral sources have increased assimilation and retention compared with sulfate mineral sources [32,41]. Equally noted by Harvey et al. [72], liver concentrations are frequently used as the standard for assessing mineral status in livestock, nonetheless, they should not be used as an absolute indicator of trace mineral status [73]. In turn, Harvey et al. [72] reported greater hepatic mRNA expression of metallothionein 1A (MT) in AAC supplemented cows, and similar hepatic mRNA expression of Cu-transporter protein (Cutting), and Cu-Zn-superoxide dismutase one (SOD) compared with INR cows at calving (Table 2). These genes are associated with Cu and Zn metabolism in the liver and provide boosted insight into the Zn and Cu status of cattle [74,75,76,77]. Hence, these outcomes advise that Cu and Zn status between AAC and INR cows may not take differed during gestation and upon calving as denoted by differences observed in liver concentrations of these trace minerals.
To complement and provide further biological back up to Marques et al. [59], Harvey et al. [72] also examined trace mineral concentrations in placental cotyledons and calf liver at nativity and 24 h afterward birth. These authors, however, reported no differences in trace mineral contour of placental and calf liver samples from AAC-supplemented cows or those receiving INR diets (Table three). Supplementing gestating beef cows with AAC also did not impact hepatic mRNA expression of Cutting, MT, or SOD in calves at birth or 24 h after (Tabular array 3), further suggesting similar hepatic Cu and Zn metabolism during early life [75,77,78]. Harvey et al. [72] also reported liver concentrations of Cu, Co, Mn, and Zn decreased in all calves from birth to 24 h after birth (Table 3), which was expected given that the bovine neonate depends heavily on liver stores of trace minerals for postnatal utilization due to low concentrations of these elements in colostrum and milk [37]. Appropriately, hepatic mRNA expression of Cut and MT increased in all calves 24 h after birth (Tabular array 3), over again reflecting the greater activity in hepatic tissue and utilization of trace minerals [37,79].
In an effort to elucidate the physiological mechanisms by which providing the AAC diet to gestating beefiness cows stimulated offspring growth via programming effects reported past Marques et al. [59], Harvey et al. [72] evaluated mRNA expression of genes associated with adipogenic and myogenic activities in the longissimus muscle (LM) in calves at nascence and at weaning. Supplementing gestating beef cows with Cu, Co, Mn, and Zn as organic complexed instead of sulfate sources did not alter mRNA expression of genes associated with these activities in the LM of calves at birth or upon weaning, despite the established role of trace minerals on musculus development and adipogenesis [80,81]. These outcomes resulted in similar offspring growth from nativity until weaning (Tabular array 4). Harvey et al. [72] besides reported like milk product and profile between AAC and INR cows (Table 4), indicating that whatever potential treatment effects on offspring responses were not caused by alterations in milk yield during early on lactation [72]. Secretion of trace elements in the milk is strictly regulated within the mammary gland, avoiding imbalances even when trace minerals are supplied in excess of requirements [82], corroborating the lack of handling differences reported by Harvey et al. [72] for lactation variables. Marques et al. [59] besides did not written report differences in offspring weaning responses betwixt AAC and INR supplemented cows, despite a 13 kg numerical increase in weaning BW from AAC cows. Collectively, Harvey et al. [72] reported that supplementing Co, Cu, Zn, and Mn equally organic complexed or sulfate sources yielded similar cow-calf productive responses until weaning.
Impacts on Female person Progeny Reared as Replacement Heifers
Both male and female person offspring were reared for slaughter past Marques et al. [59], whereas maternal diet and weaning BW have tremendous impacts on female reproductive development [27,59], and enquiry investigating trace mineral supplementation during gestation and female progeny performance is limited. For example, no differences in puberty attainment or pregnancy status were reported for heifers built-in to organically supplemented dams compared to inorganic supplemented cohorts [83]. However, Price et al. [83] began supplementation approximately 82 d prior to calving, well subsequently ovarian evolution of female progeny was complete [84]. Therefore, Harvey et al. [85] aimed to evaluate the furnishings of supplementing organic complexed or sulfate sources of Co, Cu, Mn, and Zn to beef cows during the second and third trimesters of gestation on mail-weaning responses of the female offspring reared as replacement heifers.
In Harvey et al. [85], heifers built-in to AAC-supplemented dams reached puberty before in the experiment compared with INR cohorts (Figure i). Puberty onset is highly influenced by body composition and evolution [86], whereas attainment and age at puberty differed between heifers from AAC and INR cows despite similar BW gain between treatments (Table v). In turn, heifers from INR cows had greater mRNA expression of myogenin and tended to accept greater mRNA expression of paired box gene 7 (PAX7) in the LM compared with AAC cohorts (Table 5). Myogenin is a regulatory factor in the LM musculus that influences postnatal growth through differentiation and fusion of satellite cells with existing muscle fibers [five,87], whereas PAX7 is necessary for satellite cell specification and survival [88,89]. Hence, Harvey et al. [85] speculated that heifers built-in to INR supplemented cows had a greater population of satellite cells undergoing differentiation at the time of sampling. Further, no treatment differences were detected in the LM for genes associated with adipogenic activities (Table 5), however, the intramuscular region is typically the last depot for adipose tissue to be deposited in the growing creature [90] and the puberty process is more than likely influenced by subcutaneous fat accretion [91,92,93]. Myogenic factors are downregulated equally cattle mature and muscle fibers are fully developed [5,94]; therefore Harvey et al. [85] noted that treatment differences noted in the LM for mRNA expression of myogenin and PAX7 may be indicative of accelerated physiological maturation in heifers born to AAC-supplemented cows [95].
The role of trace mineral source on reproductive performance is complex, especially given the plethora of processes each element participates in, and the essentiality of each in all aspects of evolution [32]. Previous research demonstrated lactating beef cows supplemented with organic Co, Cu, Mn, and Zn yielded a greater number of culturable oocytes and transferable embryos compared with cohorts receiving sulfate sources of the same elements [47]. Primordial germs cells within the developing ovary actively utilise machinery and enzymes against reactive oxygen species (ROS) to maintain prison cell integrity [96], and increased antioxidant enzyme action and decreased ROS production during germ cell development results with trace mineral supplementation [97,98]. Moreover, trace mineral deficiency, specifically Zn, leads to epigenetic defects and impaired ovarian development during the fetal period [99]. Thus, it became plausible to speculate that supplementing AAC to gestating cows favored the ovarian development of heifer offspring, protecting ovarian cells and follicles from endogenous ROS during development compared to heifers born to INR-supplemented cows [85].
5. Conclusions
Nutritional manipulation during periods of developmental plasticity such as the embryonic, fetal, and neonatal periods exert lasting effects on skeletal muscle and adipose tissue development, health, and overall performance of offspring [3]. Identifying specific nutritional strategies targeting these periods of development provides a unique opportunity to optimize the efficiency and profitability of beef production systems. Supplementing beef cows with organic complexed sources of Co, Cu, Mn, and Zn during gestation instead of no supplementation, enhanced passage of Zn and Cu from maternal to fetal tissues, resulting in life-long programming effects on offspring productivity and wellness. Further, heifers born to cows receiving organic-complexed trace minerals exhibited hastened puberty attainment compared with cohorts from cows that received sulfate sources. These outcomes were not associated with trace mineral deficiencies in either scenario, but rather the programmatic effects of additional Cu, Co, Mn, and Zn intake past cows. Results from these experiments are novel, and suggest that supplementing organic-complexed Co, Cu, Mn, and Zn to gestating beefiness cows enhances offspring growth and evolution, leading to long-term effects on performance, health, and accelerated reproductive development of female progeny.
Writer Contributions
Conceptualization, Yard.M.H., R.F.C. and R.d.Southward.Grand.; writing—original typhoon preparation, K.M.H., R.F.C. and R.d.Southward.M.; writing—review and editing, Thousand.Grand.H., R.F.C. and R.d.S.M.; supervision, R.F.C. All authors have read and agreed to the published version of the manuscript.
Funding
This inquiry received no external funding.
Institutional Review Board Argument
Non applicative.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicative.
Acknowledgments
The authors thank collaborating faculty, staff, and students that have helped conduct research in our laboratory and described in this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- 2050 High-Level Experts Forum: The Forum. Available online: http://world wide web.fao.org/wsfs/forum2050/wsfs-forum/en/ (accessed on 11 February 2021).
- Robinson, D.L.; Cafe, L.M.; Greenwood, P.L. Meat science and musculus biological science symposium: Developmental programming in cattle: Consequences for growth, efficiency, carcass, muscle, and beef quality characteristics. J. Anim. Sci. 2013, 91, 1428–1442. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.West.; Wallace, J.M.; Spencer, T.E. BOARD-INVITED REVIEW: Intrauterine growth retardation: Implications for the animal sciences. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef]
- Patel, M.Due south.; Srinivasan, M. Metabolic programming in the immediate postnatal life. Ann. Nurt. Metab. 2011, 58, xviii–28. [Google Scholar] [CrossRef]
- Du, M.; Tong, J.; Zhao, J.; Underwood, K.R.; Zhu, M.; Ford, South.P.; Nathanielsz, P.W. Fetal programming of skeletal musculus development in ruminant animals. J. Anim. Sci. 2010, 88, E51–E60. [Google Scholar] [CrossRef] [PubMed]
- Carcass beef grades and standards|Agricultural Marketing Service. Bachelor online: https://www.ams.usda.gov/grades-standards/carcass-beef-grades-and-standards (accessed on 12 February 2021).
- Roberts, A.J.; Petersen, 1000.K.; Funston, R.North. BEEF SPECIES SYMPOSIUM: Tin can we build the cowherd by increasing longevity of females? J. Anim. Sci. 2015, 93, 4235–4243. [Google Scholar] [CrossRef] [PubMed]
- Ireland, J.J.; Smith, M.Westward.; Scheetz, D.; Jimenez-Krassel, F.; Folger, J.K.; Ireland, J.Fifty.H.; Mossa, F.; Lonergan, P.; Evans, A.C.O. Does size affair in females? An Overview of the impact of the high variation in the ovarian reserve on ovarian function and fertility, utility of anti-Müllerian hormone every bit a diagnostic marker for fertility and causes of variation in the ovarian reserve in cattle. Reprod. Fertil. Dev. 2010, 23, i–14. [Google Scholar] [CrossRef]
- Sullivan, T.M.; Micke, G.C.; Greer, R.M.; Irving-Rodgers, H.F.; Rodgers, R.J.; Perry, V.E.A. Dietary manipulation of Bos indicus × heifers during gestation affects the reproductive development of their heifer calves. Reprod. Fertil. Dev. 2009, 21, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.; Clark, P.K. Fetal undernutrition and disease in later life. Rev. Reprod. 1997, 2, 105–112. [Google Scholar] [CrossRef]
- Bong, A.Due west. Prenatal programming of postnatal productivity and health of livestock: A cursory review. Aust. J. Exp. Agric. 2006, 46, 725–732. [Google Scholar] [CrossRef]
- Robinson, J.J.; Sinclair, Grand.D.; McEvoy, T.Grand. Nutritional effects on foetal growth. Anim. Sci. J. 1999, 68, 315–331. [Google Scholar] [CrossRef]
- Funston, R.N.; Larson, D.M.; Vonnahme, Thousand.A. Effects of maternal nutrition on conceptus growth and offspring performance: Implications for beef cattle production. J. Anim. Sci. 2010, 88, E205–E215. [Google Scholar] [CrossRef]
- Zhu, M.-J.; Ford, S.P.; Nathanielsz, P.West.; Du, Thousand. Effect of maternal food brake in sheep on the development of fetal skeletal muscle. Biol. Reprod. 2004, 71, 1968–1973. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhu, M.-J.; Dodson, Chiliad.V.; Du, M. Developmental Programming of fetal skeletal muscle and adipose tissue evolution. J. Genom. 2013, 1, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Zhu, M.J.; Underwood, K.R.; Hess, B.W.; Ford, S.P.; Du, M. AMP-activated protein kinase and adipogenesis in sheep fetal skeletal musculus and 3T3-L1 cells. J. Anim. Sci. 2008, 86, 1296–1305. [Google Scholar] [CrossRef]
- Yan, X.; Huang, Y.; Zhao, J.-X.; Long, Due north.Grand.; Uthlaut, A.B.; Zhu, 1000.-J.; Ford, South.P.; Nathanielsz, P.W.; Du, M. Maternal obesity-impaired insulin signaling in sheep and induced lipid accumulation and fibrosis in skeletal muscle of offspring. Biol. Reprod. 2011, 85, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Perry, One thousand.A.; Cushman, R. Effect of age at puberty/conception date on cow longevity. Vet. Clin. Northward. Am. Food Anim. 2013, 29, 579–590. [Google Scholar] [CrossRef]
- Martin, J.L.; Vonnahme, K.A.; Adams, D.C.; Lardy, G.P.; Funston, R.N. Effects of dam nutrition on growth and reproductive performance of heifer calves. J. Anim. Sci. 2007, 85, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Mossa, F.; Kenny, D.; Jimenez-Krassel, F.; Smith, G.W.; Berry, D.; Butler, Due south.; Fair, T.; Lonergan, P.; Ireland, J.J.; Evans, A.C.O. Undernutrition of heifers during the first trimester of pregnancy diminishes size of the ovarian reserve in female person offspring. Biol. Reprod. 2009, 81, 135. [Google Scholar] [CrossRef]
- Mossa, F.; Carter, F.; Walsh, S.Westward.; Kenny, D.A.; Smith, One thousand.W.; Ireland, J.50.H.; Hildebrandt, T.B.; Lonergan, P.; Ireland, J.J.; Evans, A.C.O. Maternal undernutrition in cows impairs ovarian and cardiovascular systems in their offspring. Biol. Reprod. 2013, 88. [Google Scholar] [CrossRef]
- Ireland, J.50.H.; Scheetz, D.; Jimenez-Krassel, F.; Themmen, A.P.N.; Ward, F.; Lonergan, P.; Smith, G.Due west.; Perez, Thousand.I.; Evans, A.C.O.; Ireland, J.J. Antral follicle count reliably predicts number of morphologically good for you oocytes and follicles in ovaries of young adult cattle. Biol. Reprod. 2008, 79, 1219–1225. [Google Scholar] [CrossRef]
- Cushman, R.A.; Allan, M.F.; Kuehn, L.A.; Snelling, W.M.; Cupp, A.S.; Freetly, H.C. Evaluation of antral follicle count and ovarian morphology in crossbred beef cows: Investigation of influence of stage of the estrous cycle, age, and birth weight. J. Anim. Sci. 2009, 87, 1971–1980. [Google Scholar] [CrossRef] [PubMed]
- Mossa, F.; Walsh, Due south.W.; Butler, S.T.; Berry, D.P.; Carter, F.; Lonergan, P.; Smith, Chiliad.West.; Ireland, J.J.; Evans, A.C.O. Depression numbers of ovarian follicles ≥ 3 mm in diameter are associated with low fertility in dairy cows. J. Dairy Sci. 2012, 95, 2355–2361. [Google Scholar] [CrossRef]
- O'Neil, Thou.M.; Westward, Southward.M.; Maia, T.S.; Cardoso, R.C.; Williams, G.L. Effects of maternal nutrition on secretion of leptin in the neonatal heifer and interaction of maternal and postnatal diet on age at puberty and postpubertal secretion of luteinizing hormone. J. Anim. Sci. 2019, 97, 140–141. [Google Scholar] [CrossRef]
- Larson, D.K.; Martin, J.50.; Adams, D.C.; Funston, R.N. Wintertime grazing system and supplementation during late gestation influence performance of beef cows and steer progeny. J. Anim. Sci. 2009, 87, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Cushman, R.A.; Perry, G.A. Developmental programming of fertility in livestock. Vet. Clin. North. Am. Food Anim. 2019, 35, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.; Chidzanja, S.; Kind, Thousand.; Lok, F.; Owens, P.; Owens, J. Placental control of fetal growth. Reprod. Fertil. Dev. 1995, 7, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Bohnert, D.Due west.; Stalker, Fifty.A.; Mills, R.R.; Nyman, A.; Falck, South.J.; Cooke, R.F. Late gestation supplementation of beef cows differing in torso condition score: Effects on cow and calf performance. J. Anim. Sci. 2013, 91, 5485–5491. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.B.; Faulkner, D.B.; Shike, D.W. Influence of prepartum dietary energy on beef cow performance and calf growth and carcass characteristics. Livest. Sci. 2016, 184, 21–27. [Google Scholar] [CrossRef]
- Radunz, A.E.; Fluharty, F.50.; Relling, A.E.; Felix, T.L.; Shoup, L.Thou.; Zerby, H.Northward.; Loerch, S.C. Prepartum dietary energy source fed to beef cows: Two. Effects on progeny postnatal growth, glucose tolerance, and carcass composition. J. Anim. Sci. 2012, 90, 4962–4974. [Google Scholar] [CrossRef]
- Hostetler, C.East.; Kincaid, R.L.; Mirando, 1000.A. The part of essential trace elements in embryonic and fetal development in livestock. Vet. J. 2003, 166, 125–139. [Google Scholar] [CrossRef]
- Spears, J.W. Minerals in Forages. In Forage Quality, Evaluation, and Utilization; John Wiley & Sons, Ltd.: Hoboken, NJ, The states, 1994; pp. 281–317. ISBN 978-0-89118-579-6. [Google Scholar]
- McDowell, L.R. Feeding minerals to cattle on pasture. Anim. Feed Sci. Technol. 1996, 60, 247–271. [Google Scholar] [CrossRef]
- Greene, L.W. Designing mineral supplementation of fodder programs for beef cattle. J. Anim. Sci. 2000, 77, 1–9. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine. Nutrient Requirements of Beef Cattle: Eighth Revised Edition; The National Academies Press: Washington, DC, USA, 2016; ISBN 978-0-309-31702-3. [Google Scholar]
- Underwood, E.J.; Suttle, N.F. The mineral Nutrition of Livestock, third ed.; CABI Publishing: Wallingford, Oxon, Uk, 1999; ISBN 978-0-85199-128-3. [Google Scholar]
- Hidiroglou, 1000.; Knipfel, J.E. Maternal-fetal relationships of copper, manganese, and sulfur in ruminants. A review. J. Dairy Sci. 1981, 64, 1637–1647. [Google Scholar] [CrossRef]
- Gooneratne, S.R.; Christensen, D.A. A survey of maternal copper status and fetal tissue copper concentrations in Saskatchewan bovine. Tin. J. Anim. Sci. 2011, 69, 141–150. [Google Scholar] [CrossRef]
- Hansard, Southward.L. Physiological Behavior of Manganese in Gravid Cattle, Sheep and Swine. In Proc. Isotope Studies on the Physiology of Domestic Animals; International Atomic Energy Agency: Vienna, Austria, 1972. [Google Scholar]
- Spears, J.W. Organic trace minerals in ruminant diet. Anim. Feed Sci. Technol. 1996, 58, 151–163. [Google Scholar] [CrossRef]
- Brown, T.F.; Zeringue, L.1000. laboratory evaluations of solubility and structural integrity of complexed and chelated trace mineral supplements. J. Dairy Sci. 1994, 77, 181–189. [Google Scholar] [CrossRef]
- Nocek, J.Due east.; Socha, One thousand.T.; Tomlinson, D.J. The effect of trace mineral fortification level and source on functioning of dairy cattle. J. Dairy Sci. 2006, 89, 2679–2693. [Google Scholar] [CrossRef]
- Kellogg, D.Due west.; Socha, M.T.; Tomlinson, D.J.; Johnson, A.B. Effects of feeding cobalt glucoheptonate and metallic specific amino acid complexes of zinc, manganese, and copper on lactation and reproductive operation of dairy cows. Prof. Anim. Sci. 2003, 19, i–9. [Google Scholar] [CrossRef]
- Griffiths, L.M.; Loeffler, S.H.; Socha, M.T.; Tomlinson, D.J.; Johnson, A.B. Furnishings of supplementing complexed zinc, manganese, copper and cobalt on lactation and reproductive performance of intensively grazed lactating dairy cattle on the due south isle of New Zealand. Anim. Feed Sci. Technol. 2007, 137, 69–83. [Google Scholar] [CrossRef]
- Uchida, G.; Mandebvu, P.; Ballard, C.Due south.; Sniffen, C.J.; Carter, M.P. Effect of feeding a combination of zinc, manganese and copper amino acid complexes, and cobalt glucoheptonate on operation of early lactation high producing dairy cows. Anim. Feed Sci. Technol. 2001, 93, 193–203. [Google Scholar] [CrossRef]
- Dantas, F.G.; Reese, South.T.; Filho, R.5.O.; Carvalho, R.S.; Franco, M.A.; Abbott, C.R.; Payton, R.R.; Edwards, J.50.; Russell, J.R.; Smith, J.Thousand.; et al. Issue of complexed trace minerals on cumulus-oocyte complex recovery and in vitro embryo production in beef cattle. J. Anim. Sci. 2019, 97, 1478–1490. [Google Scholar] [CrossRef] [PubMed]
- Rabiee, A.R.; Lean, I.J.; Stevenson, Grand.A.; Socha, G.T. Effects of feeding organic trace minerals on milk production and reproductive functioning in lactating dairy cows: A meta-analysis. J. Dairy Sci. 2010, 93, 4239–4251. [Google Scholar] [CrossRef]
- Ahola, J.1000.; Baker, D.S.; Burns, P.D.; Mortimer, R.Yard.; Enns, R.M.; Whittier, J.C.; Geary, T.W.; Engle, T.E. Effect of copper, zinc, and manganese supplementation and source on reproduction, mineral condition, and performance in grazing beef cattle over a two-year period. J. Anim. Sci. 2004, 82, 2375–2383. [Google Scholar] [CrossRef] [PubMed]
- Osorio, J.S.; Trevisi, Due east.; Li, C.; Drackley, J.K.; Socha, 1000.T.; Loor, J.J. Supplementing Zn, Mn, and Cu from amino acid complexes and Co from cobalt glucoheptonate during the peripartal menses benefits postpartal cow performance and claret neutrophil function. J. Dairy Sci. 2016, 99, 1868–1883. [Google Scholar] [CrossRef]
- Batistel, F.; Osorio, J.South.; Ferrari, A.; Trevisi, Due east.; Socha, M.T.; Loor, J.J. Immunometabolic status during the peripartum period is enhanced with supplemental Zn, Mn, and Cu from Amino Acid complexes and Co from Co glucoheptonate. PLoS I 2016, 11, e0155804. [Google Scholar] [CrossRef]
- Overton, T.R.; Yasui, T. Applied applications of trace minerals for dairy cattle. J. Anim. Sci. 2014, 92, 416–426. [Google Scholar] [CrossRef]
- Abuelo, A.; Hernández, J.; Benedito, J.L.; Castillo, C. The importance of the oxidative condition of dairy cattle in the periparturient period: Revisiting antioxidant supplementation. J. Anim. Physiol. Anim. Nutr. 2015, 99, 1003–1016. [Google Scholar] [CrossRef]
- Roshanzamir, H.; Rezaei, J.; Fazaeli, H. Colostrum and milk functioning, and claret amnesty indices and minerals of Holstein cows receiving organic Mn, Zn and Cu sources. Anim. Nutr. 2020, 6, 61–68. [Google Scholar] [CrossRef]
- Formigoni, A.; Fustini, Grand.; Archetti, L.; Emanuele, S.; Sniffen, C.; Biagi, G. Effects of an organic source of copper, manganese and zinc on dairy cattle productive performance, wellness status and fertility. Anim. Feed Sci. Technol. 2011, 164, 191–198. [Google Scholar] [CrossRef]
- Van Emon, Yard.; Sanford, C.; McCoski, S. Impacts of bovine trace mineral supplementation on maternal and offspring product and wellness. Animals 2020, x, 2404. [Google Scholar] [CrossRef] [PubMed]
- Weiss, W.P.; Colenbrander, V.F.; Cunningham, M.D.; Callahan, C.J. Selenium/vitamin E: Role in disease prevention and weight gain of neonatal calves. J. Dairy Sci. 1983, 66, 1101–1107. [Google Scholar] [CrossRef]
- Pepper, M.R.; Black, M.K. B12 in fetal development. Semin. Cell Dev. Biol. 2011, 22, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Marques, R.S.; Cooke, R.F.; Rodrigues, 1000.C.; Cappellozza, B.I.; Mills, R.R.; Larson, C.K.; Moriel, P.; Bohnert, D.W. Effects of organic or inorganic cobalt, copper, manganese, and zinc supplementation to late-gestating beefiness cows on productive and physiological responses of the offspring. J. Anim. Sci. 2016, 94, 1215–1226. [Google Scholar] [CrossRef] [PubMed]
- Food Requirements of Beef Cattle: Seventh Revised Edition; The National Academies Printing: Washington, DC, USA, 2000.
- Stanton, T.L.; Whittier, J.C.; Geary, T.W.; Kimberling, C.V.; Johnson, A.B. Furnishings of trace mineral supplementation on cow-calf performance, reproduction, and immune function. Prof. Anim. Sci. 2000, 16, 121–127. [Google Scholar] [CrossRef]
- Arthington, J.D.; Swensont, C.Grand. Effects of trace mineral source and feeding method on the productivity of grazing Braford cows. Prof. Anim. Sci. 2004, 20, 155–161. [Google Scholar] [CrossRef]
- Kegley, E.B.; Brawl, J.J.; Beck, P.A. Bill E. Kunkle Interdisciplinary Beefiness Symposium: Touch of mineral and vitamin status on beefiness cattle immune function and health. J. Anim. Sci. 2016, 94, 5401–5413. [Google Scholar] [CrossRef]
- McClure, S.J. How minerals may influence the development and expression of amnesty to endoparasites in livestock. Parasite Immunol. 2008, 30, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Wintergerst, Eastward.S.; Maggini, S.; Hornig, D.H. Contribution of selected vitamins and trace elements to immune part. Ann. Nutr. Metab. 2007, 51, 301–323. [Google Scholar] [CrossRef] [PubMed]
- Jacometo, C.B.; Osorio, J.Due south.; Socha, M.; Corrêa, M.N.; Piccioli-Cappelli, F.; Trevisi, E.; Loor, J.J. Maternal consumption of organic trace minerals alters calf systemic and neutrophil mRNA and MicroRNA indicators of inflammation and oxidative stress. J. Dairy Sci. 2015, 98, 7717–7729. [Google Scholar] [CrossRef]
- Reynolds, L.P.; Vonnahme, K.A. Livestock as models for developmental programming. Anim. Forepart. 2017, vii, 12–17. [Google Scholar] [CrossRef]
- Caton, J.South.; Crouse, M.Due south.; Reynolds, 50.P.; Neville, T.L.; Dahlen, C.R.; Ward, A.One thousand.; Swanson, G.C. Maternal nutrition and programming of offspring energy requirements. Transl. Anim. Sci. 2019, 3, 976–990. [Google Scholar] [CrossRef] [PubMed]
- Ashworth, C.J.; Antipatis, C. Micronutrient programming of development throughout gestation. Reproduction 2001, 122, 527–535. [Google Scholar] [CrossRef]
- Du, M.; Ford, S.P.; Zhu, M.-J. Optimizing livestock production efficiency through maternal nutritional management and fetal developmental programming. Anim. Front. 2017, vii, v–11. [Google Scholar] [CrossRef]
- USDA APHIS|NAHMS Beef Cow-Calf Studies. Available online: https://www.aphis.usda.gov/aphis/ourfocus/animalhealth/monitoring-and-surveillance/nahms/nahms_beef_cowcalf_studies (accessed on 12 February 2021).
- Harvey, K.M.; Cooke, R.F.; Colombo, E.A.; Rett, B.; Sousa, O.A.; Harvey, L.Thousand.; Russell, J.R.; Pohler, M.G.; Brandão, A.P. Supplementing organic-complexed or inorganic Co, Cu, Mn, and Zn to beefiness cows during gestation: Physiological and productive responses of cows and their offspring until weaning. J. Anim. Sci. 2021. [Google Scholar] [CrossRef]
- McDowell, L.R. Minerals in Animal and Man Nutrition, 2nd ed.; Elsevier: Amsterdam, Holland, 2003; ISBN 978-0-444-51367-0. [Google Scholar]
- Sturtz, L.A.; Diekert, K.; Jensen, Fifty.T.; Lill, R.; Culotta, V.C. A Fraction of yeast Cu,Zn-superoxide dismutase and its metallochaperone, CCS, localize to the intermembrane space of mitochondria: A physiological role for SOD1 in guarding against mitochondrial oxidative harm*. J. Biol. Chem. 2001, 276, 38084–38089. [Google Scholar] [CrossRef] [PubMed]
- Coyle, P.; Philcox, J.C.; Carey, L.C.; Rofe, A.Yard. Metallothionein: The multipurpose protein. Cell. Mol. Life Sci. 2002, 59, 627–647. [Google Scholar] [CrossRef]
- Prohaska, J.R.; Gybina, A.A. Intracellular copper transport in mammals. J. Nutr. 2004, 134, 1003–1006. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Archibeque, Due south.Fifty.; Engle, T.E. Label and identification of hepatic mRNA related to copper metabolism and homeostasis in cattle. Biol. Trace Elem. Res. 2009, 129, 130–136. [Google Scholar] [CrossRef]
- Hansen, Due south.50.; Ashwell, M.Southward.; Legleiter, Fifty.R.; Fry, R.S.; Lloyd, K.E.; Spears, J.W. The addition of high manganese to a copper-deficient nutrition further depresses copper status and growth of cattle. Br. J. Nutr. 2008, 101, 1068–1078. [Google Scholar] [CrossRef]
- López-Alonso, Chiliad.; Prieto, F.; Miranda, 1000.; Castillo, C.; Hernández, J.; Benedito, J.50. The office of metallothionein and zinc in hepatic copper accumulation in cattle. Vet. J. 2005, 169, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, Yard.; Nagata, Y.; Wada, E.; Zammit, P.S.; Shiozuka, M.; Matsuda, R. Zinc promotes proliferation and activation of myogenic cells via the PI3K/Akt and ERK signaling cascade. Exp. Cell Res. 2015, 333, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Wei, Due south.; Zhang, L.; Zhou, 10.; Du, M.; Jiang, Z.; Hausman, G.J.; Bergen, Due west.G.; Zan, L.; Dodson, 1000.5. Emerging roles of zinc finger proteins in regulating adipogenesis. Prison cell. Mol. Life Sci. 2013, 70, 4569–4584. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B. Trace chemical element send in the mammary gland. Annu. Rev. Nutr. 2007, 27, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Toll, D.M.; O'Neil, 1000.K.; Watson, W.B., III; Due west, R.; Rae, D.O.; Irsik, D.M.; Hersom, G.J.; Yelich, J.V. 1262 Consequence of pre- and postnatal trace mineral (TM) source on angus and Brangus heifer growth and reproductive performance. J. Anim. Sci. 2016, 94, 609. [Google Scholar] [CrossRef]
- Nilsson, E.E.; Skinner, M.K. Progesterone regulation of primordial follicle assembly in bovine fetal ovaries. Mol. Cell. Endocrinol. 2009, 313, 9–16. [Google Scholar] [CrossRef]
- Harvey, 1000.M.; Cooke, R.F.; Colombo, E.A.; Rett, B.; Sousa, O.A.; Harvey, Fifty.1000.; Russell, J.R.; Pohler, K.G.; Brandão, A.P. Supplementing organic-complexed or inorganic Co, Cu, Mn, and Zn to beef cows during gestation:mail-weaning responses of offspring reared as replacement heifers or feeder cattle. J. Anim. Sci. 2021. [Google Scholar] [CrossRef]
- Schillo, K.K.; Hall, J.B.; Hileman, Due south.M. effects of diet and season on the onset of puberty in the beef heifer. J. Anim. Sci. 1992, lxx, 3994–4005. [Google Scholar] [CrossRef]
- Le Grand, F.; Rudnicki, Yard.A. Skeletal muscle satellite cells and developed myogenesis. Curr. Opin. Cell Biol. 2007, 19, 628–633. [Google Scholar] [CrossRef]
- Seale, P.; Sabourin, L.A.; Girgis-Gabardo, A.; Mansouri, A.; Gruss, P.; Rudnicki, M.A. Pax7 is required for the specification of myogenic satellite cells. Cell 2000, 102, 777–786. [Google Scholar] [CrossRef]
- Li, J.; Gonzalez, J.1000.; Walker, D.K.; Hersom, M.J.; Ealy, A.D.; Johnson, Due south.E. Evidence of heterogeneity within bovine satellite cells isolated from young and adult animals. J. Anim. Sci. 2011, 89, 1751–1757. [Google Scholar] [CrossRef]
- De Oliveira, I.M.; Paulino, P.V.R.; Marcondes, One thousand.I.; de Valadares Filho, S.C.; Detmann, E.; Cavali, J.; de Duarte, M.Due south.; Mezzomo, R. Blueprint of tissue deposition, gain and body composition of Nellore, F1 Simmental × Nellore and F1 Angus × Nellore steers fed at maintenance or advertizement libitum with two levels of concentrate in the diet. Rev. Bras. Zootec. 2011, 40, 2886–2893. [Google Scholar] [CrossRef]
- Lammoglia, 1000.A.; Bellows, R.A.; Grings, Eastward.East.; Bergman, J.W.; Bellows, Southward.Due east.; Short, R.East.; Hallford, D.Thou.; Randel, R.D. Furnishings of dietary fatty and sire brood on puberty, weight, and reproductive traits of F1 beef heifers. J. Anim. Sci. 2000, 78, 2244–2252. [Google Scholar] [CrossRef]
- Garcia, M.R.; Amstalden, M.; Williams, S.W.; Stanko, R.L.; Morrison, C.D.; Keisler, D.H.; Nizielski, Southward.E.; Williams, 1000.L. Serum leptin and its adipose gene expression during pubertal development, the estrous bike, and unlike seasons in cattle. J. Anim. Sci. 2002, 80, 2158–2167. [Google Scholar] [CrossRef]
- Roberts, A.J.; da Silva, A.G.; Summers, A.F.; Geary, T.W.; Funston, R.N. Developmental and reproductive characteristics of beef heifers classified by pubertal status at time of commencement convenance. J. Anim. Sci. 2017, 95, 5629–5636. [Google Scholar] [CrossRef] [PubMed]
- Picard, B.; Lefaucheur, L.; Berri, C.; Duclos, M.J. Muscle fibre ontogenesis in subcontract brute species. Reprod. Nutr. Dev. 2002, 42, 415–431. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.Fifty.; Busse, N.I.; Waits, C.M.; Johnson, S.East. Satellite cells and their regulation in livestock. J. Anim. Sci. 2020, 98, skaa081. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Otsuka, Chiliad.; Ebina, 1000.; Igarashi, K.; Takehara, A.; Matsumoto, M.; Kanai, A.; Igarashi, K.; Soga, T.; Matsui, Y. Distinct requirements for energy metabolism in mouse primordial germ cells and their reprogramming to embryonic germ cells. Proc. Natl. Acad. Sci. Us 2017, 114, 8289–8294. [Google Scholar] [CrossRef]
- Özkaya, M.O.; Nazıroğlu, G. Multivitamin and mineral supplementation modulates oxidative stress and antioxidant vitamin levels in serum and follicular fluid of women undergoing in vitro fertilization. Fertil. Steril. 2010, 94, 2465–2466. [Google Scholar] [CrossRef] [PubMed]
- Shi, Fifty.; Yue, Westward.; Zhang, C.; Ren, Y.; Zhu, 10.; Wang, Q.; Shi, Fifty.; Lei, F. effects of maternal and dietary selenium (Se-enriched yeast) on oxidative status in testis and apoptosis of germ cells during spermatogenesis of their offspring in goats. Anim. Reprod. Sci. 2010, 119, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Hurley, 50.; Swell, C. Fetal and neonatal development in relation to maternal trace element nutrition: Manganese, zinc, and copper. In Vitamins and Minerals in Pregnancy and Lactation; Berger, H., Ed.; Raven Printing: New York, NY, Us, 1988; Book 16, pp. 215–230. [Google Scholar]
Figure one. Puberty attainment of replacement heifers from beefiness cows that received diets containing supplemental sulfate sources of Co, Cu, Mn, and Zn (INR) or organic-complexed source of Co, Cu, Mn, and Zn (AAC) during gestation. Adapted from Harvey et al. [85]. Heifers were weaned on d 367 of the experiment, at ~vii mo of age. A treatment × day interaction was detected (p < 0.01). Within days: † 0.05 ≤ p ≤ 0.x; * p ≤ 0.05; ** p < 0.01.
Effigy i. Puberty attainment of replacement heifers from beef cows that received diets containing supplemental sulfate sources of Co, Cu, Mn, and Zn (INR) or organic-complexed source of Co, Cu, Mn, and Zn (AAC) during gestation. Adapted from Harvey et al. [85]. Heifers were weaned on d 367 of the experiment, at ~7 mo of age. A treatment × day interaction was detected (p < 0.01). Inside days: † 0.05 ≤ p ≤ 0.ten; * p ≤ 0.05; ** p < 0.01.
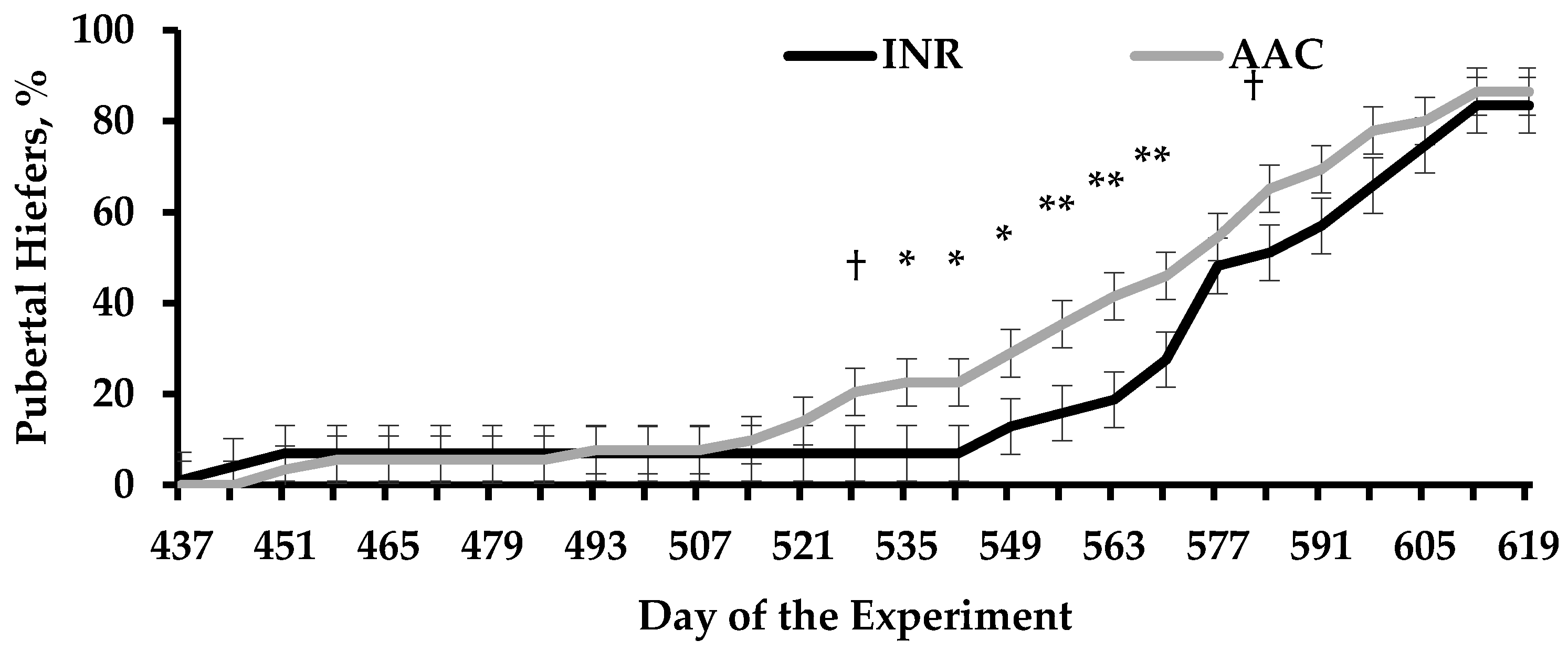
Table 1. Physiological, wellness, and productive responses from beef cows (and their offspring) that received diets containing no supplemental Cu, Co, Mn, and Zn (CON); sulfate sources of Cu, Co, Mn, and Zn (INR); or organic complexed source of Cu, Mn, Co, and Zn (AAC) during the terminal trimester of gestation i.
Tabular array 1. Physiological, health, and productive responses from beef cows (and their offspring) that received diets containing no supplemental Cu, Co, Mn, and Zn (CON); sulfate sources of Cu, Co, Mn, and Zn (INR); or organic complexed source of Cu, Mn, Co, and Zn (AAC) during the last trimester of gestation ane.
| Item | CON | INR | AAC |
|---|---|---|---|
| Cow liver 2, mg/kg | |||
| Co | 0.21 a | 0.40 b | 0.44 c |
| Cu | 69 a | 155 b | 129 c |
| Mn | viii.7 | 9.0 | 8.7 |
| Zn | 211 a | 230 b | 235 b |
| Cotyledon 3, mg/kg | |||
| Co | 0.13 a | 0.20 b | 0.24 b |
| Cu | 3.88 a | 4.75 a,b | 5.12 b |
| Mn | 22.0 | 18.2 | 22.nine |
| Zn | 65 | 66 | 68 |
| Calf liver 3, mg/kg | |||
| Co | 0.09 a | 0.12 b | 0.xiii b |
| Cu | 362 a | 428 a,b | 450 b |
| Mn | 5.82 | 5.22 | 5.83 |
| Zn | 456 a | 562 a,b | 660 b |
| Dogie birth BW, kg | 42.1 | 41.half-dozen | 40.8 |
| Calf weaning BW, kg | 212 a | 223 a,b | 236 b |
| Treated for BRD symptoms 4, % | 42.3 a | 59.1 a | 20.0 b |
| BW prior to slaughter, kg | 649 a | 663 a,b | 680 b |
| Hot carcass weight, kg | 409 a | 418 a,b | 428 b |
Table 2. Concentrations of Co, Cu, Mn, and Zn and expression of genes in the liver in beefiness cows that received diets containing supplemental sulfate sources of Co, Cu, Mn, and Zn (INR) or organic-complexed source of Co, Cu, Mn, and Zn (AAC) during gestation 1,two.
Tabular array 2. Concentrations of Co, Cu, Mn, and Zn and expression of genes in the liver in beef cows that received diets containing supplemental sulfate sources of Co, Cu, Mn, and Zn (INR) or organic-complexed source of Co, Cu, Mn, and Zn (AAC) during gestation one,2.
| Item | INR | AAC |
|---|---|---|
| Liver mineral profile | ||
| Co, mg/kg | ||
| Late gestation | 0.59 b | 0.68 a |
| Calving | 0.58 | 0.55 |
| Cu, mg/kg | ||
| Late gestation | 125 a | 81.9 b |
| Calving | 118 a | 42.ix b |
| Mn, mg/kg | ||
| Late gestation | ten.v | ten.0 |
| Calving | 12.0 | 10.6 |
| Zn, mg/kg | ||
| Late gestation | 307 | 341 |
| Calving | 173 a | 129 b |
| Gene mRNA expression 3 | ||
| Cu-transporter protein | ||
| Late gestation | 1.67 | one.70 |
| Calving | one.95 | 1.75 |
| Metallothionein 1A | ||
| Tardily gestation | 26.1 | 22.8 |
| Calving | 36.4 b | 65.6 a |
| Superoxide dismutase ane | ||
| Late gestation | 1.99 | two.eleven |
| Calving | 2.02 | two.04 |
Table 3. Concentrations of Co, Cu, Mn, and Zn and expression of genes in the liver in beef cows that received diets containing supplemental sulfate sources of Co, Cu, Mn, and Zn (INR) or organic-complexed source of Co, Cu, Mn, and Zn (AAC) during gestation ane,ii.
Table 3. Concentrations of Co, Cu, Mn, and Zn and expression of genes in the liver in beef cows that received diets containing supplemental sulfate sources of Co, Cu, Mn, and Zn (INR) or organic-complexed source of Co, Cu, Mn, and Zn (AAC) during gestation 1,2.
| Item | INR | AAC |
|---|---|---|
| Liver mineral profile | ||
| Co, mg/kg | ||
| Cotyledon | 0.494 | 0.533 |
| Dogie birth | 0.202 | 0.188 |
| Calf 24 h later on nascency | 0.153 | 0.148 |
| Cu, mg/kg | ||
| Cotyledon | 8.38 | 9.07 |
| Calf birth | 394 | 399 |
| Calf 24 h afterward nascency | 303 | 291 |
| Mn, mg/kg | ||
| Cotyledon | 18.9 | 18.iv |
| Calf nascence | 5.96 | 6.06 |
| Dogie 24 h after nativity | 4.lxx | 4.83 |
| Zn, mg/kg | ||
| Cotyledon | 90.9 | 93.4 |
| Calf birth | 823 | 869 |
| Dogie 24 h later nascency | 676 | 637 |
| Factor mRNA expression 3 | ||
| Cu-transporter protein | ||
| Nascency | 2.19 | 2.23 |
| 24 h after birth | 2.51 | ii.53 |
| Metallothionein 1A | ||
| Nascence | 33.7 | 32.9 |
| 24 h after nascency | 59.iv | 68.four |
| Superoxide dismutase 1 | ||
| Birth | 2.92 | 2.96 |
| 24 h afterwards nativity | 2.77 | 2.70 |
Table 4. Lactation responses of beef cows that received diets containing supplemental sulfate sources of Co, Cu, Mn, and Zn (INR) or organic-complexed Co, Cu, Mn, and Zn (AAC) during gestation, and performances responses of their offspring until weaning one.
Tabular array 4. Lactation responses of beef cows that received diets containing supplemental sulfate sources of Co, Cu, Mn, and Zn (INR) or organic-complexed Co, Cu, Mn, and Zn (AAC) during gestation, and performances responses of their offspring until weaning 1.
| Item | INR | AAC |
|---|---|---|
| Lactation responses | ||
| Milk yield, kg/d | vi.62 | 6.98 |
| Milk mineral profile | ||
| Co, ppm | ||
| Colostrum | 0.0052 | 0.0055 |
| Milk | 0.00030 | 0.00035 |
| Cu, ppm | ||
| Colostrum | 0.985 | 0.997 |
| Milk | 0.049 | 0.062 |
| Mn, ppm | ||
| Colostrum | 0.061 | 0.055 |
| Milk | 0.014 | 0.016 |
| Zn, ppm | ||
| Colostrum | 15.3 | xiii.7 |
| Milk | 0.014 | 0.016 |
| Calf operation responses | ||
| Calf birth BW, kg | 30.5 | 30.eight |
| Dogie weaning BW, kg | 183 | 178 |
Table v. Productive responses and mRNA expression of longissimus muscle genes in replacement heifers built-in from beef cows that received diets containing supplemental sulfate sources of Co, Cu, Mn, and Zn (INR) or organic complexed source of Co, Cu, Mn, and Zn (AAC) during gestation ane.
Table 5. Productive responses and mRNA expression of longissimus musculus genes in replacement heifers born from beef cows that received diets containing supplemental sulfate sources of Co, Cu, Mn, and Zn (INR) or organic complexed source of Co, Cu, Mn, and Zn (AAC) during gestation one.
| Particular | INR | AAC |
|---|---|---|
| Initial body weight 2, kg | 202 | 197 |
| Concluding body weight 2, kg | 332 | 326 |
| Average daily gain ii, kg/d | 0.618 | 0.604 |
| Final puberty attainment 3, % | 83.five | 86.4 |
| Age at puberty, d | 418 a | 399 b |
| Body weight at puberty, kg | 319 | 310 |
| mRNA expression 4 | ||
| Adipocyte fatty acid bounden protein | 4.68 | v.10 |
| Myogenin | four.59 a | 2.87 b |
| Paired box cistron vii | i.91 x | 1.70 y |
| Peroxisome proliferator-activated receptor-γ | 1.62 | 1.53 |
| Publisher's Annotation: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access commodity distributed under the terms and weather of the Artistic Commons Attribution (CC Past) license (https://creativecommons.org/licenses/by/four.0/).
Source: https://www.mdpi.com/2076-2615/11/4/1159/htm
0 Response to "Impact of Mineral and Vitamin Status on Beef Cattle Immune Function and Health"
Post a Comment